
Глава 2
Биоритмы и ритмы космоса
2.1. Ритмозадающие свойства космогеофизических факторов
Эффекты воздействия космогеофизических факторов на биосферу очень разнообразны. Так, скорость окисления тимоловых соединений в зависимости от солнечной активности является показателем биотропности рассматриваемых параметров. Установлена корреляция между временем полуокисления унитиола (SH-групп) и числами Вольфа, потоком радиоизлучения Солнца на частоте 202 МГц и днями хромосферных вспышек (Соколовский В. В., 1982а; 1982б; 1984). Выявлен феномен флуктуаций в водных растворах белков и других химических веществ. Флуктуации при измерениях электрофоретической подвижности белков и фликкерных шумов сходны в синхронных опытах (Шноль С. Э. с соавт., 1983). Выделены различные гармоники макроскопических флуктуаций водных растворов белков и других веществ (Удальцова Н. В. с соавт., 1983). Обнаружена связь выводимых кортикостероидов с ГМП с разной направленностью параметров (Загорская Е. А. с соавт., 1982). Для усредненных индивидуальных хронограмм экспрессии рецепторов Т-лимфоцитов в течение дня обнаружены достоверные корреляции с характером гравитационного поля на структурно-функциональную периодичность клеток (Гариб Ф. Ю. с соавт., 1995).
Ритм является правильным чередованием лидерства двух противоположностей, являющихся залогом качественной устойчивости, условием равновесия протовоположных тенденций через управление неравновесными состояниями (Степанова С. И., 1977; Мартынюк В. С., 1996). Он понимается как устойчивое, закономерно проявляющееся повторение в организации систем и процессов и служит характеристикой функциональных систем и фактором естественного отбора, может быть подвижен и пластичен с определенным диапазоном вариаций, закрепленных филогенетически с динамически-стереотипной организацией. Филогенетически закреплено одно из свойств организма, заключающееся в «опережающем отражении» (Анохин П. К., 1979), смысл которого состоит в том, что биосистемы научились из совокупной комбинации внешних факторов делать необходимый «полезный для себя вывод» по временной организации своего собственного многомерного биологического времени. Довольно часто в природе получается так, что максимумы и минимумы космических и гелиогеофизических явлений совпадают с таковыми в органическом мире (Чижевский А. Л., 1976; Чернышев В. Б. с соавт., 1984). Следовательно между ними существуют определенные временные соотношения — биоритмы на клеточном, органном и целостном уровне: суточные, недельные, месячные, сезонные, годовые.
Клетка является самостоятельной функциональной единицей. Жизнедеятельность организма состоит взаимодействия клеток. В них происходят процессы анаболизма и катаболизма в различных соотношениях, что и составляет ее основные ритмы (Малахов Г. П., 1994). В принципе возможна синхронизация любым периодическим физическим агентом при условии близости воздействующих частот к собственным частотам биосистемы (Блехман И. И., 1971). Скорее всего, имеют место их естественное ранжирование, сочетания и комбинации, играющие роль многофакторности и многокомпонентности биоритмов. Одно из ведущих мест в системе внешних естественно-природных факторов играет ГМП, особенно в диапазоне сверхнизкочастотных колебаний (5 Гц), зависящих от СА. На центральную нервную систему эти параметры влияют особенно значимо (Сидякин В. Г. с соавт., 1983; 1996б). Изоляция и экранирование от естественного фона ЭМП приводят к десинхронозам или необратимым изменениям (Wever R., 1968; Рыжиков Г. В. с соавт., 1982). Макроскопические флуктуации в природе возрастают с увеличением гелиогеомагнитной активности, при экранировании от ЭМП не нарушаются, а искусственные электростатические поля также сохраняют их ход, но с изменением амплитуд отдельных реакций (Шноль С. Э. с соавт., 1983).
К микробиоритмам относят ритм электроэнцефалограммы человека. В пределах мезо — и макродиапазонов временные структуры обладают гармоническим единством: очень многие периоды и флуктуации обнаруживают соизмеримость и целочисленную кратность, относительно близким периодам присуще постоянство соотношений между фазами. В древности это свойство называли «музыкой сфер» (Чечельницкий А. М., 1980).
Комплексное исследование метеорологических и гелиогеофизических факторов является наиболее плодотворным при изучении биоритмов (Malek J. et al., 1962; Агаджанян Н. А., 1967; Орехов К. В. с соавт., 1981; Андронова Т. И. с соавт., 1982; Агаджанян Н. А. с соавт., 1985). В ходе многих процессов наблюдаются одни и те же периоды, что и при регистрации в ГМП: в метеорологических, акустических, инфразвуковых шумах, радиоактивности атмосферы, концентрации аэроионов, ЭМП во всем диапазоне сверхнизких частот. Но таких исследований практически нет (Опалинская А. М., Агулова Л. П., 1984).
2.2. Механизмы биоритмологических закономерностей
ПеМП могут выступать «датчиком времени» при эндогенных и экзогенных десинхронозах, способны изменять временную организацию физиологических систем. Характер и направленность изменений зависят от физиологического состояния организма и сопровождаются рассогласованием или нормализацией биоритмов (Темурьянц Н. А., Шехотихин А. В., 1995). Суточные ритмы и соотношения гормональной системы: гипофиз — надпочечники, половые гормоны, соматотропный и тиреоидные гормоны, инсулин, содержание глюкозы, рениновая активность плазмы крови, альдостерон, вазопрессин, окситоцин, паратгормон, кальцитонин — обнаружены и в норме, и в патологии (Дедов И. И., Дедов В. И., 1992). В результате действия естественных ЭМП развивается совокупность компенсаторно-приспособительных факторов (Макеев В. Б., Темурьянц Н. А., 1982; Темурьянц Н. А., 1982). Моделирование таких влияний невозможно, в силу того что естественные ЭМП и лабораторные сильно отличаются по спектру (Опалинская А. М., Агулова Л. П., 1984).
Синхронизация — распространенное явление в природе (Блехман И. И., 1981). При определенной амплитуде внешнего сигнала — вынуждающей силы — происходит захват частоты автоколебательной системы в небольшой, относительно узкой полосе частот синхронизации. Захват может происходить на гармонических частотах в целое число раз больших или меньших частоты вынуждающего сигнала. Около полосы синхронизации частот возможно появление особого колебательного режима — биений. В таком случае нет постоянного соотношения между фазой внешнего воздействия и автоколебательной фазой биосистемы (Корнетов А. Н. с соавт., 1988). Существует определенное соотношение самосопряженных частот в системе в ответ на внешнее воздействие, при котором внешняя вынуждающая частота разлагается на произведение частот по правилу «золотого сечения» (Дегтярев Г. М. с соавт., 1991).
Таким образом, в исследованиях с воздействием на флуктуирующие колебательные биосистемы могут также наблюдаться захват частоты, синхронизация, биения или резонанс. Вблизи границ полосы захвата наблюдались биения (Адамчук А. С., 1972). Существуют предположения о делении биоэффектов на 3 основных типа: аддитивный, антагонистический, синергический (Опалинская А. М., Агулова Л. П., 1984). В ходе флуктуаций биологических параметров организм постоянно претерпевает адаптационные сдвиги. Адаптация — это необходимое условие существования живого, выражение диалектического единства организма с внешней средой (Новиков В. С., Деряпа Н. Р., 1992). Эта проблема многоплановая. Часто физиологическую адаптацию связывают с гомеостазом и физиологическими механизмами, определяющими устойчивость систем организма (Сапов И. А., Новиков В. С., 1984). Такой взгляд развивал и В. И. Медведев (1984) с точки зрения системной реакции человеческого организма и системной приспособляемости к конкретным условиям с учетом генной фенотипической нормы реакции. Некоторые авторы процесс адаптации характеризуют непрерывностью и периодичностью (Агаджанян Н. А., 1972; Казначеев В. П., 1980). Позднее процесс адаптации дополнился условием оптимальности ко множеству природных и социальных факторов и стал рассматриваться в качестве признака здоровья (Агаджанян Н. А., 1982).
Для биологических колебательных систем вынуждающей силой может быть любой периодически изменяющийся внешний фактор. Деление ритмов на эндогенные и экзогенные очень условно и отражает скорее степень их изученности, чем реальную причину колебаний. Не очень удачным представляется также деление ритмов на диапазоны с приставкой circa (около) (Ашофф Ю., 1984). Cуществует деление биоритмов по спектру частот: микроритмы, мезоритмы, макроритмы, периоды большой длительности. Одним из внешних водителей ритмов этой периодичности являются секторные границы межпланетного МП (Оль А. И., 1973). Выделяют периоды большой длительности — ритмика крупномасштабных экологических изменений, регулярные климатические колебания в десятки тысяч лет, длительные вариации СА, КЛ, ГМП, глобальные изменения среды обитания катастрофического характера (Митчел Дж. М. мл. с соавт., 1982; Владимирский Б. М., Кисловский Л. Д., 1985; Прудников И. М. с соавт., 1996).
Ритмоупорядоченность генетически детерминирована с наследованием датчиков времени внешней среды и соотношений гармоник определенного диапазона. В качестве синхронизирующего агента живая материя использует естественные флуктуации ГМП, при перепадах значения которого возникают различные варианты десинхронозов, проявляющиеся в виде изменений ритмов сна (Моисеева Н. И., Сысуев В. М., 1980). Приведенные факты свидетельствуют, что колебания в биологических и биофизических и химических процессах являются не случайными, а упорядоченными во времени под влиянием факторов внешней среды. Совпадение биоритмов с периодами гелиомагнитной активности — признак синхронизации биоритмов с параметрами внешней среды (Brown F. A., Ir., 1965; Владимирский Б. М., 1982). Таким образом, и наши результаты наглядно показали причинную обусловленность многолетних, сезонных и многомесячных синергетических флуктуаций и вариаций ферментных, биохимических, гематологических, нейрофизиологических и психологических показателей организма космическими излучениями, солнечной активностью, ионосферными процессами в совокупности с ГМП и акустико-гравитационными воздействиями.
2.3. Спектрально-частотные особенности биопроцессов и космогеофизических факторов
За последние 3–4 десятилетия изучены флуктуации многих физико-химических и биологических процессов. Выяснились особенности многих видов флуктуаций, в том числе фликкерных шумов — типа 1/f, где f — частота процесса. Под шумом в узком смысле понимают белый шум, характеризующийся тем, что его амплитудный спектр распределен по нормальному закону, а спектральная плотность мощности постоянна для всех частот. Фликкерный шум характеризуется наиболее низкочастотным спектром по сравнению с шумами иных видов и для оценки стационарности требует более длительного интервала наблюдения по сравнению с прочими частотами (Денда В., 1993). Одна из главных закономерностей флуктуаций заключается в зависимости амплитуды события от его встречаемости (закон 1/f) и неоднозначности — синхронные колебания одинакового типа могут быть разного знака и разной амплитуды. Флуктуирующий объект — сложная система, биосфера Земли — четкая синхронизированная иерархия систем. Особенности закономерностей связаны с результатами совокупных релаксаций напряжений системы, накопленных за достаточно большое время от возмущений разного масштаба, перераспределяющихся по внутренним связям. Даже случайные толчки приводят не к хаотическим изменениям, а к статистическим переключениям подпрограмм системных согласованных релаксаций (Опалинская А. М., Агулова Л. П., 1984). Такие заключения дают возможность прогноза и управления целостными системами (Шноль С. Э. с соавт., 1983). В нашей работе эти положения получили полнейшие подтверждения в силу того, что основные исследованные космогелиофизические факторы — КЛ и СА — имеют примерно 11 — и 22-летнюю цикличность и выявление их трендов требует длительного наблюдения, а соответствующие проявления биологических параметров и систем должны быть по крайней мере не менее продолжительными. Нами получены результаты относительно спектральных гармоник и соответствующих им периодов, свидетельствующие о совпадении трендов среднемесячных биохимических, электролитных, ферментных, гематологических систем и отдельных их параметров с основными среднемесячными показателями космогелиогеофизических процессов. Определенные сведения о корреляциях выявлены в отношении дыхательной и сердечно-сосудистой систем, нейрофизиологических процессов головного мозга и психического состояния здоровых людей. Сведения научной литературы подтверждают наши наблюдения.
В настоящее время существует много публикаций о различных моделях, описывающих условия возникновения и поведение флуктуаций в физических, химических, биологических системах: о неравновесных системах с флуктуациями (Николис Г., Пригожин И., 1979), колебаниях разнородных систем (Кешнер М. С., 1982). Предполагается существование фундаментального закона природы, приложимого ко всем неравновесным системам и проявляющегося в шуме типа 1/f. К шуму 1/f были отнесены флуктуации ферментативной активности титра SH-групп и некоторых других характеристик водных растворов белков и небелковых веществ (Удальцова Н. В., 1982). Распределение скоростей реакций (спектры), в которых наблюдаются макроскопические флуктуации, полимодально. К шуму 1/f относят изменения многих космических процессов, ОЧСП (Timashev S. F., Kostioutchenco I. G., 1995; Tanizuka N. et al., 1995), сопротивления водных растворов ионов, диффузионные потоки ионов в мембранных каналах. К этому классу относят многие процессы биологии, химии, экономики (Богданов А. А., 1928; Гапонов-Грехов А. В., Рабинович М. И., 1980; Козьменко С. Ю., 1995).
Ниже, на рис. 2.1. и 2.2., приводятся 2 примера спектра-графика, близких по типу к 1/f для космического и биологического параметра. Спектральные плотности вычислены с применением спектрального окна Парзена. Кратко суть процедуры сводится к тому, что квантуется (дискретизируется) любой физический или биологический процесс так, что он представляет собой данные Xt, t = 1, 2,…, N, отсчитанные через промежутки времени d. Далее вычисляется сглаженная выборочная оценка нормированной спектральной плотности по специальным формулам (Бокс Дж., Дженкинс Г., 1974).
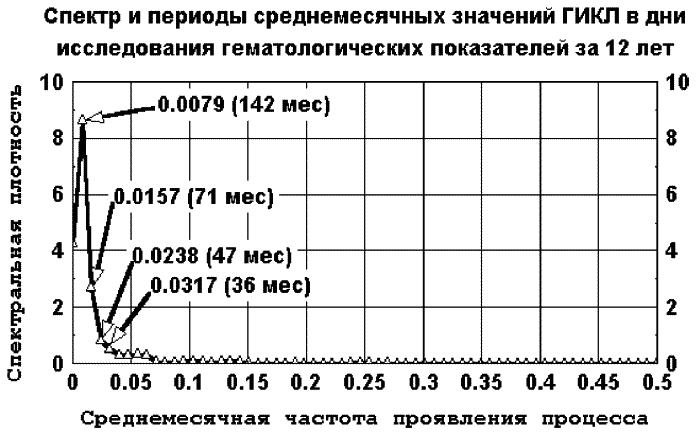
Рис. 2.1. 1/f-подобный спектр и периоды среднемесячных значений ГИКЛ в дни исследования гематологических параметров в период с 1977 по 1988 г. Здесь частота f соответствует около 11-летнему циклу (142 мес.)

Рис. 2.2. Спектр и периоды среднемесячных значений содержания палочкоядерных нейтрофилов в циркулирующей крови в утренние часы в период с 1977 по 1988 г. наглядно демонстрируют, что долгосрочные периоды и спектры совпадают с показателями ГИКЛ и, следовательно, имеют 1/f — подобные закономерности
Биологический ритм, от клеточного уровня до уровня поведения, в подавляющих случаях подчиняется флуктуациям по закону 1/fn, где n — ближайшая частота (или ранг частот) устройства. Возможны 3 варианта причин: ионный перенос флуктуаций клеточных мембран, регулирующих ионные потоки в клетке в полуинтервалах, полное управление неврогенной природы или суперпозиция случайных событий. Величины биологических параметров всегда флуктуируют во времени. Существуют доказательства 1/f-подобных флуктуаций биологических параметров от клеточного до поведенческого уровня. Возможны несколько механизмов генерации 1/f-подобных биологических ритмических флуктуаций Во-первых, 1/f-ионные флуктуационные потоки мембран клеток модулируют поток ионов внутрь клеток, которые в свою очередь модулируются изменениями интервалов импульсов клеток и нервов. Исследован новый механизм функциональной регуляции ионной проводимости каналов в зависимости от флуктуации окружающей среды (Bezrukov S. M. et al., 1995; Pustovoit M. A. et al., 1995). Во-вторых, временная задержка и ответы системной нервной регуляции могут быть причиной 1/f-модуляций, например флуктуации сердечного ритма и кровяного давления (Musha T., Yamamoto M., 1995).
Изучены 1/f-флуктуации нейронной активности нервной системы во время регистрации быстрых движений глаз кошек в период сна. Этот феномен наблюдался в обширной области мозга, такой как ретикулярная формация, таламус, церебральный кортекс, гиппокамп. Предполагают существование глобальной модулирующей системы в мозге с участием серотониноэргетики и холиноэргетики (Yamamoto M., 1995). Развитие автономной нервной системы плода выглядит так, что большую роль играют 1/f-и 1/f2-спектры сердцебиений плода, варьирующиеся на частотах ниже 0,05 Гц, которые поддерживают отношения парасимпатической и/или симпатической нервных систем и описывают процесс роста и взросления (Shono H. et al., 1995).
Экспериментальные данные указывают на присутствие в спектре интервалов сердцебиений в частотах 10–4 — 10–5 Гц. Каждый интервал сердечных сокращений имеет тенденцию быть некоррелированным с другими наблюдениями в течение 3 — 24 часов. Это может быть причиной 1/f-спектрального профиля хотя величина спектральной плотности для частот ниже 10–5 Гц не значима; наибольшие значения спектра указывают на ультранизкие частоты, что указывает на включение интервалов сердечных сокращений в общие автокорреляции в течение длительного времени. Авторы приходят к выводу о необходимости проверки таких гипотез в течение многомесячных наблюдений (Yamamoto M. et al., 1995).
В то же время есть указания на внешюю причинность таких вариаций. Изучение динамики сердечного ритма — R-R-интервала человека в зависимости от геофизических и метеорологических условий выявило корреляции индекса централизации и амплитуды респираторной волны кардиоритма с атмосферным давлением и геомагнитной активностью подтверждает такие предположения (Smirnova N. A., Augustinaite E. E., 1995). Исследование статистических свойств сердечного ритма на основе клинических данных о сердечной патологии показало возрастание абсолютной величины 1/fn у кардиальных больных (Ulbikas J. et al., 1995). В связи с этим корреляции показателей дыхательной и сердечно-сосудистой систем с ионосферными параметрами, приводимые ниже в настоящей работе, выглядят совершенно естественными.












